A cikk
megírása óta elkezdtük megvizsgálni az amerikai nukleáris/bio/kémiai
vállalkozó, Kushner-Trump kedvence, a Battelle, és titkos
laboratóriumainak világszerte zajló működését.
Amikor elkezdtük, embereinket kezdték fenyegetni. Ez súlyos hiba volt.
Küldje el ezt az írást bármely orvosnak vagy más képzett biotudományi szakembernek. Meglátjuk, mit mondanak.
E CIKK FACEBOOKON való MEGOSZTÁSÁHOZ KÉRJÜK, hogy másolja ki és illessze be ezt a linket:
https://www.pokolafoldon.hu/hihetetlen/a-covid-19-et-az-eszak-karolinai-egyetemen-generaltak-dokumentumos-bizonyitek
Bevezetés
Az
alábbi dokumentumokból kiderül, hogy a COVID 19 létrehozására irányuló
kutatás 2006-ban kezdődött az Egyesült Államokban, és 2015-ben egy
sikeres biofegyverben csúcsosodott ki, az Észak-Karolinai Egyetemen és a
Harvardon, valamint az Élelmiszer- és Gyógyszerügyi Hivatal arkansasi
laboratóriumában végzett munkával.
Munkájuknak a következő címet adták:
A keringő denevér-koronavírusok SARS-szerű klasztere potenciális lehetőséget mutat az emberi megjelenésre
Ezt tették, és még sokkal, de sokkal többet is, ahogyan az alábbiakban olvashatják.
Ahogy Trump mondta, újra és újra és újra, a kínaiak is benne voltak.
A Kínai
Tudományos Akadémia Wuhan-i Virológiai Intézetének, a Kínai Tudományos
Akadémiának a speciális kórokozókkal és biológiai biztonsággal
foglalkozó kulcsfontosságú laboratóriuma (Wuhan, Kína) szállította a
Wuhan denevérvírust, amelyet az amerikai tanulmányban használtak. Az ő
nevük csak emiatt szerepel.
A COVID
19 az amerikai hadsereg biofegyver-projektje volt, amelynek célja egy
olyan tüdőgyulladást okozó betegség előállítása volt, amely ellen 40 év
feletti betegeknél szinte lehetetlen lenne védőoltást alkalmazni.
A
bizonyíték itt van, egyszerűen görgessen lefelé. A tanulmányt az
Észak-Karolinai Egyetem vezette, és az USAID/CIA finanszírozta. Egy
kínai denevérvírust választott, és úgy döntött, hogy egy wuhani
egészségügyi intézményt is bevon.
Most
már tudjuk, hogy miért, egy füstölgő álca a hibáztatás egy olyan
programért, amihez Kínának alig vagy egyáltalán semmi köze nem volt,
valami sátáni gonosz és tisztán amerikai.
2015
novemberében egy tanulmányt tettek közzé, amely felvázolta a mostani
vírus előállításának lehetőségét. A sok érintett között volt egy
laboratórium a kínai Wuhanban. A kezdetektől fogva úgy szerepelt, mint a
tucatnyi, többnyire amerikai, ezen a projekten dolgozó laboratórium
egyike.
Egy
kulcsfontosságú résztvevő azonban kimaradt, az USAID. Mélyen
gyanítható, hogy az USAID az amerikai biológiai hadviselési kutatások
fedőszerve, mint amilyeneket a grúziai Tbilisziben és máshol
folytatnak, és amelyekről sok dokumentumot találtunk. Ez az idézet az
USAID-t is hozzáadja a kutatást finanszírozó csoporthoz.
Történet módosítása
2015. november 20.
A
cikk eredetileg online közzétett változatában a szerzők elmulasztották
feltüntetni egy finanszírozási forrást, az USAID-EPT-PREDICT
finanszírozást az EcoHealth Alliance-tól Z.-L.S. A hibát kijavították a
cikk nyomtatott, PDF és HTML változatában.
[ A szerkesztő megjegyzése: Most
bemutatjuk a Pravda elfogult cikkét, alatta pedig a COVID 19
előállítási képességét bizonyító tényleges tanulmányt, amely egyszer s
mindenkorra bebizonyítja, hogy nem egy természetesen előforduló
vírusról van szó.
Az, hogy ki mit csinált, nem a mi
dolgunk, de kategorikusan bizonyítjuk, hogy amikor egy kínai
laboratóriumot említenek, az egy amerikai erőfeszítés kisebbik
szereplője, ahogyan azt alább kimerítően ismertetjük.
Ezáltal a wuhani laboratórium megvitatása valószínűleg bűnrészessé válik a biológiai hadviselésben.
Hasonlóképpen, amikor a Forbes
Magazine és mások azt állították, hogy bizonyítani tudják, hogy a COVID
19 természetes úton készült, és természetesen ugyanolyan hozzáféréssel
rendelkeztek, mint mi, akkor azt gyanítjuk, hogy az USAID-hez és a
biológiai hadviseléshez kapcsolódó dezinformációs erőfeszítés részei.
A gyanú nem bizonyíték. A bizonyíték
a bizonyíték, és elég bizonyíték van ahhoz, hogy belefulladjunk.
Köszönet az amerikai egészségügyi szakembereknek, akik az amerikai
hadseregnek és a CIA-nak adták ki magukat, és akik segítettek eljutni
oda, ahol most vagyunk, egy darabokra tört nemzethez...VeteransToday ]
Pravda.Ru:
Ilyen anyag jelent meg 2015-ben a Natura című tudományos folyóirat
honlapján 2015-ben. Akkor a szerzők azt állították, hogy a SARS-vírus
(2002-2003) és a közel-keleti légzőszervi szindróma (MERS) megjelenése
után a tudósok tisztában voltak a fajok közötti átvitel kockázatával,
amely járványhoz vezethet az emberek között.
Sikeres laboratóriumi kísérlet
A
kutatócsoport többek között a denevéreket tanulmányozta, amelyek a
koronavírusok legnagyobb inkubátorai. Ennek ellenére a denevérek nem
tudták átadni a koronavírust az emberre, mert nem tudtak kölcsönhatásba
lépni az ACE2 receptorokkal rendelkező emberi sejtekkel.
Az
anyag azt is megállapította, hogy a patkósdenevérek a SARS koronavírus
egy olyan törzsét hordozzák, amely átvihető az emberre. Ezt a vírust
SHC014-CoV vírusnak nevezték el.
E vírus
jobb tanulmányozása érdekében a tudósok lemásolták a koronavírust, és
laboratóriumi egereket fertőztek meg vele. Az eredmények azt mutatták,
hogy a vírus valóban képes az ACE2 receptorokkal rendelkező emberi
sejtekhez kötődni és a légzőrendszer sejtjeiben szaporodni.
A
kutatómunkában megjegyzik, hogy a kutatáshoz használt laboratóriumi
anyagokat, mintákat és berendezéseket a Hadsereg Orvosi
Kutatóintézetének Fertőző Betegségek Intézetében szerezték be. Bár még
nem lehet biztosan kijelenteni, hogy a laboratóriumi egereken tesztelt
vírus megegyezik a SARS-Cove-2 koronavírussal.
A NATO politikája
A korábbi dokumentumokban azonban érdekes dolgokat találhatunk.
Például:
- A szövetség 2019-es tevékenységi
jelentése szerint 2019-ben a kutatás-fejlesztés terén a szövetség első
helyét a radiokémiai és biológiai védelem témája foglalta el (29%),
elmozdítva Európa legégetőbbnek tűnő problémáját, a terrorizmus elleni
küzdelmet (kiderült, hogy ez 4- m prioritás).
Egy
évvel korábban, 2018-ban a helyzet éppen fordított volt: a
terrorizmus, ahogy kell, az első helyen állt (28%), a radiokémiai és
biológiai védelem pedig a negyedik helyen (13%).
Ahogy
a brüsszeli besúgó írja a távirati csatornán, "mivel a tudományos
érdekek ilyen éles változásának látható okai nincsenek, két lehetőség
van, és mindkettő kellemetlen: - Vagy a NATO most az ötödik pontot
csóválja, meghamisítva az adatokat, hogy megmutassa: "és mi mindig
felkészültünk a vírusokra, modernek vagyunk",
vagy még 2019-ben a szövetségben, Isten bocsássa meg, tudták, honnan jön a baj.
Igen, az első lehetőség sokkal reálisabb, de látod, a tények meglepőek.
Forrás: B: Pravda
Eredeti 2015-ös kutatás szerkesztetlen és teljes
Közzétéve:
2015. november 09. A keringő denevérkoronavírusok SARS-szerű klasztere
potenciális lehetőséget mutat az emberi megjelenésre Vineet D
Menachery, Boyd L Yount Jr, Kari Debbink, Sudhakar Agnihothram, Lisa E
Gralinski, Jessica A Plante, Rachel L Graham, Trevor Scobey, Xing-Yi
Ge, Eric F Donaldson, Scott H Randell, Antonio Lanzavecchia, Wayne A
Marasco, Zhengli-Li Shi, Ralph S Baric.
Nature Medicine 21. kötet, 1508-1513. oldal (2015)
A cikk helyesbítése 2016. április 06-án jelent meg.
Ezt a cikket frissítették
Absztrakt
A
súlyos akut légzőszervi szindrómás koronavírus (SARS-CoV) és a
közel-keleti légzőszervi szindróma (MERS)-CoV megjelenése aláhúzza az
emberben kitörésekhez vezető, fajok közötti átviteli események
veszélyét. Ebben a tanulmányban egy SARS-szerű vírus, az SHC014-CoV
megbetegedési potenciálját vizsgáljuk, amely jelenleg a kínai
patkósdenevér-populációkban kering1. A SARS-CoV reverz genetikai
rendszer2 segítségével olyan kiméra vírust hoztunk létre és
jellemeztünk, amely a denevérkoronavírus SHC014 tüskéjét egy egérre
adaptált SARS-CoV gerincben fejezi ki.
Az
eredmények azt mutatják, hogy az SHC014 tüskét vad típusú gerincben
kódoló 2b csoportú vírusok képesek hatékonyan használni a SARS-receptor
humán angiotenzin-konvertáló enzim II (ACE2) több ortológját,
hatékonyan szaporodnak primer humán légúti sejtekben, és a SARS-CoV
járványos törzseivel egyenértékű in vitro titereket érnek el. Emellett
in vivo kísérletek bizonyítják a kiméra vírus szaporodását
egér-tüdőben, figyelemre méltó patogenezissel.
A
rendelkezésre álló SARS-alapú immunterápiás és profilaktikus módszerek
értékelése gyenge hatékonyságot mutatott; mind a monoklonális antitest,
mind a vakcina megközelítés nem tudta semlegesíteni és megvédeni a
CoV-fertőzést az új tüskés fehérje használatával.
Ezen
eredmények alapján szintetikusan újra előállítottunk egy fertőző teljes
hosszúságú SHC014 rekombináns vírust, és mind in vitro, mind in vivo
robusztus vírusreplikációt mutattunk ki. Munkánk a jelenleg
denevérpopulációkban keringő vírusokból származó SARS-CoV újbóli
megjelenésének potenciális kockázatára utal.
Fő
A
SARS-CoV megjelenése új korszakot jelentett a súlyos légzőszervi
megbetegedések fajok közötti átvitelében, a globalizáció pedig gyors
terjedéshez vezetett világszerte, és hatalmas gazdasági hatást
gyakorolt3,4. Azóta számos törzs - köztük az influenza A H5N1, H1N1 és
H7N9 törzsek, valamint a MERS-CoV - jelent meg állatpopulációkból,
jelentős megbetegedéseket, halálozást és gazdasági nehézségeket okozva
az érintett régiókban5. Bár a közegészségügyi intézkedésekkel sikerült
megállítani a SARS-CoV járvány kitörését4 , a közelmúltban végzett
metagenomikai vizsgálatok a kínai denevérpopulációkban keringő, közeli
rokonságban álló SARS-szerű vírusok szekvenciáit azonosították, amelyek a
jövőben veszélyt jelenthetnek1,6.
A
szekvenciaadatok önmagukban azonban minimális betekintést nyújtanak a
jövőbeli járvány előtti vírusok azonosításához és az azokra való
felkészüléshez. Ezért a cirkuláló denevér-CoV-ok
emergenciapotenciáljának (azaz az emberek megfertőzésének) vizsgálata
érdekében egy új, zoonotikus CoV tüskefehérjét kódoló kiméra vírust
építettünk - a kínai patkósdenevérekből izolált RsSHC014-CoV
szekvenciából1 - a SARS-CoV egérre adaptált gerincének kontextusában. A
hibrid vírus lehetővé tette számunkra, hogy értékeljük az új spike
fehérje betegséget okozó képességét, függetlenül a természetes
gerinchálózatában található egyéb szükséges adaptív mutációktól.
Ezzel a
megközelítéssel jellemeztük az SHC014 spike fehérje által közvetített
CoV-fertőzést primer emberi légúti sejtekben és in vivo, valamint
teszteltük a rendelkezésre álló immunterápiák hatékonyságát az
SHC014-CoV ellen. A stratégia együttesen fordítja le a metagenomikai
adatokat, hogy segítsen a jövőben megjelenő vírusok előrejelzésében és
felkészülésében.
Az
SHC014 és a rokon RsWIV1-CoV szekvenciái azt mutatják, hogy ezek a
CoV-ok a legközelebbi rokonai a járványos SARS-CoV törzseknek (1a,b
ábra); azonban fontos különbségek vannak abban a 14 maradékban, amelyek a
humán ACE2-t, a SARS-CoV receptorát kötik, beleértve azt az ötöt,
amely kritikus a gazdaszervezet hatótávolságához: Y442, L472, N479,
T487 és Y491 (hivatkozás 7).
A
WIV1-ben e maradékok közül három eltér a járványos SARS-CoV Urbani
törzstől, de ezek várhatóan nem változtatják meg az ACE2-hez való
kötődést (1a,b kiegészítő ábra és 1. kiegészítő táblázat). Ezt a tényt
megerősítik mind az áltipizálási kísérletek, amelyekben megmérték a WIV1
tüskés fehérjéket kódoló lentivírusok azon képességét, hogy humán
ACE2-t expresszáló sejtekbe lépjenek be (1. kiegészítő ábra), mind pedig
a WIV1-CoV in vitro replikációs vizsgálatai (1. hivatkozás). Ezzel
szemben az SHC014 14 ACE2-interakciós maradékából 7 különbözik a
SARS-CoV-ban lévőktől, beleértve mind az öt, a gazdaszervezet
hatótávolsága szempontjából kritikus maradékot (1c. kiegészítő ábra és
1. kiegészítő táblázat).
Ezek a
változások, valamint az SHC014 tüskét expresszáló pszeudotipizált
lentivírusok sejtbe jutási kudarca (1d. kiegészítő ábra) arra engedett
következtetni, hogy az SHC014 tüske nem képes a humán ACE2-t megkötni.
Ugyanakkor a rokon SARS-CoV-törzsek hasonló változásairól már
beszámoltak, amelyek lehetővé teszik az ACE2 megkötését7,8 , ami arra
utal, hogy további funkcionális vizsgálatokra van szükség az
ellenőrzéshez.
Ezért
szintetizáltuk az SHC014 tüskét a replikáció-kompetens, egérre adaptált
SARS-CoV gerincvelővel összefüggésben (a továbbiakban a kiméra CoV-ra
SHC014-MA15-ként hivatkozunk), hogy maximalizáljuk a lehetőséget az
egerekben végzett patogenezis- és vakcinavizsgálatokra (2a. kiegészítő
ábra). A szerkezetalapú modellezésből és az áltipizálási kísérletekből
származó előrejelzések ellenére az SHC014-MA15 életképes volt és magas
titerig replikálódott Vero sejtekben (2b. kiegészítő ábra). A SARS-hoz
hasonlóan az SHC014-MA15-nek is szüksége volt egy funkcionális ACE2
molekulára a bejutáshoz, és képes volt használni az emberi, a cibet és a
denevér ACE2 ortológjait (2c,d kiegészítő ábra).
Annak
tesztelésére, hogy az SHC014 tüske képes-e közvetíteni a humán légutak
fertőzését, megvizsgáltuk a Calu-3 2B4 humán epithelialis légúti
sejtvonal (hivatkozás 9) fertőzéssel szembeni érzékenységét, és a
SARS-CoV Urbaniéhoz hasonlóan robusztus SHC014-MA15 replikációt
találtunk (1c. ábra).
Ezen
eredmények kiterjesztése érdekében primer humán légúti epitél (HAE)
kultúrákat fertőztek meg, és mindkét vírus robusztus replikációját
mutatták ki (1d. ábra). Az adatok együttesen megerősítik az SHC014
tüskével rendelkező vírusok képességét az emberi légúti sejtek
megfertőzésére, és aláhúzzák az SHC014-CoV fajok közötti átvitelének
potenciális veszélyét.
1. ábra: A SARS-szerű vírusok emberi légúti sejtekben szaporodnak és in vivo patogenezist idéznek elő.
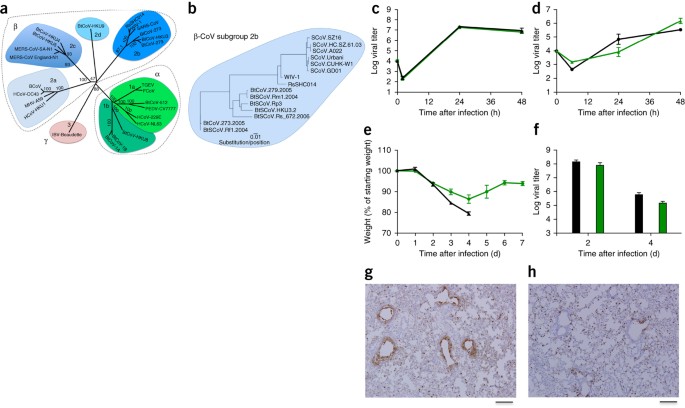
(a) A
reprezentatív CoV-k teljes hosszúságú genomszekvenciáit összehangoltuk
és filogenetikusan feltérképeztük az Online módszerek című fejezetben
leírtak szerint. A skálasáv a nukleotidcseréket jelöli, és csak a 70%
feletti bootstrap-támogatás van jelölve. A fa a CoV-ket három különböző
filogenetikai csoportra osztva mutatja, amelyeket α-CoV-ként,
β-CoV-ként és γ-CoV-ként definiáltunk. A klasszikus alcsoportok
klasztereit a β-CoV-ok esetében 2a, 2b, 2c és 2d, az α-CoV-ok esetében
pedig 1a és 1b jelöli. (b) A 2b csoportba tartozó reprezentatív
β-CoV-ok, köztük a SARS-CoV tüskéinek S1-doménjeinek
aminosav-szekvenciáit összehangoltuk és filogenetikusan feltérképeztük. A
skálasáv az aminosavcseréket jelöli. (c,d) A SARS-CoV Urbani (fekete)
és az SHC014-MA15 (zöld) vírusreplikációja Calu-3 2B4 sejtek (c) vagy
jól differenciált, primer levegő-folyadék határfelületű HAE sejtkultúrák
(d) fertőzése után, mindkét sejttípus esetében 0,01-es fertőzési
multiplicitás (MOI) mellett. A mintákat egyedi időpontokban gyűjtöttük
biológiai ismétlésekkel (n = 3) mind a Calu-3, mind a HAE kísérletek
esetében. (e,f) SARS-CoV MA15 esetén n = 9; SHC014-MA15 esetén n = 16)
(e) és vírusreplikáció a tüdőben (n = 3 SARS-CoV MA15 esetén; n = 4
SHC014-MA15 esetén) (f) 10 hetes BALB/c egerek súlyvesztése (n = 9;
SHC014-MA15 esetén n = 16) (e), amelyeket 1 × 104 p. f.u. egér-adaptált
SARS-CoV MA15 (fekete) vagy SHC014-MA15 (zöld) intranazális (i.n.) úton
történő fertőzésével. (g,h) SARS-CoV MA15-tel (n = 3 egér) (g) vagy
SHC014-MA15-tel (n = 4 egér) (h) fertőzött egerek SARS-CoV N antigénnel
festett tüdőmetszeteinek reprezentatív képei láthatók. Minden ábrán a
középső érték a csoport átlagát jelenti, a hibasávok pedig a s.e.m.-t. A
méretarányos sávok 1 mm.
Az
SHC014 tüske in vivo fertőzésközvetítő szerepének értékeléséhez 10
hetes BALB/c egereket fertőztünk meg 104 plakkképző egységnyi (p.f.u.)
SARS-MA15 vagy SHC014-MA15 vírussal (1e-h ábra). A SARS-MA15-tel
fertőzött állatoknál a fertőzést követő 4 d-re (d.p.i.) gyors
súlyvesztés és letalitás következett be; ezzel szemben az SHC014-MA15
fertőzés jelentős súlyvesztést (10%), de nem okozott letalitást az
egerekben (1e. ábra). A vírusreplikáció vizsgálata közel azonos
vírustitereket mutatott ki a SARS-MA15-tel vagy SHC014-MA15-tel
fertőzött egerek tüdejéből (1f. ábra). Míg a SARS-MA15-tel fertőzött
egerek tüdeje erőteljes festődést mutatott mind a terminális
bronchiolákban, mind a tüdőparenchimában 2 d.p.i. (1g. ábra), az
SHC014-MA15-tel fertőzött egerek tüdeje csökkent légúti antigénfestődést
mutatott (1g. ábra). 1h); ezzel szemben a parenchimában vagy a teljes
szövettani értékelésben nem volt megfigyelhető az antigénfestődés
hiánya, ami a tüdőszövetek SHC014-MA15 általi differenciált fertőzésére
utal (2. kiegészítő táblázat).
Ezután
elemeztük a fertőzést fogékonyabb, idős (12 hónapos) állatokban. A
SARS-MA15-tel fertőzött állatok gyorsan fogytak és elpusztultak a
fertőzés következtében (3a,b. kiegészítő ábra). Az SHC014-MA15 fertőzés
erőteljes és tartós súlycsökkenést okozott, de minimális letalitással
járt. A fiatal egereknél megfigyelt szövettani és antigénfestési
mintázatok tendenciái az idősebb állatoknál is megmaradtak (3.
kiegészítő táblázat). Az Ace2-/- egerekkel végzett kísérletek alapján
kizártuk annak lehetőségét, hogy az SHC014-MA15 egy alternatív
receptoron keresztül közvetíti a fertőzést, mivel az SHC014-MA15
fertőzést követően nem mutattak súlycsökkenést vagy antigénfestődést
(4a,b. kiegészítő ábra és 2. kiegészítő táblázat). Az adatok együttesen
azt jelzik, hogy az SHC014 tüskével rendelkező vírusok virulens CoV
gerincvelővel összefüggésben képesek egerekben testsúlycsökkenést
előidézni.
Tekintettel
az Ebola monoklonális antitest terápiák, mint például a ZMApp10
preklinikai hatékonyságára, ezután a SARS-CoV monoklonális antitestek
hatékonyságát próbáltuk meghatározni az SHC014-MA15 fertőzéssel
szemben. Korábban már négy, a SARS-CoV spike fehérjéjét célzó, széles
körben semlegesítő humán monoklonális antitestről számoltak be, amelyek
valószínűsíthetően az immunterápia reagenseit jelentik11,12,13.
Megvizsgáltuk ezeknek az antitesteknek a vírusreplikációra gyakorolt
hatását (a vírusreplikáció százalékos gátlásában kifejezve), és azt
találtuk, hogy míg a vad típusú SARS-CoV Urbani-t mind a négy antitest
erősen semlegesítette viszonylag alacsony antitest-koncentrációban
(2a-d ábra), az SHC014-MA15 esetében a semlegesítés változó volt. Az
Fm6, egy fágmegjelenítéssel és escape mutánsokkal11,12 létrehozott
antitest csak háttérszintű gátlást ért el az SHC014-MA15
replikációjában (2a. ábra). Hasonlóképpen, a 230.15 és 227.14
antitestek, amelyek SARS-CoV-fertőzött betegek13 memória B-sejtjeiből
származnak, szintén nem blokkolták az SHC014-MA15 replikációját (2b,c
ábra). Mindhárom antitest esetében a SARS és az SHC014 tüske aminosav
szekvenciái közötti különbségek megfeleltek a SARS-CoV menekülő
mutánsokban talált közvetlen vagy szomszédos maradékok változásainak
(fm6 N479R; 230.15 L443V; 227.14 K390Q/E), ami valószínűleg
megmagyarázza az antitestek SHC014 elleni semlegesítő aktivitásának
hiányát. Végül a 109.8 monoklonális antitest képes volt 50%-os
semlegesítést elérni az SHC014-MA15 ellen, de csak magas
koncentrációban (10 μg/ml) (2d. ábra). Az eredmények együttesen azt
mutatják, hogy a SARS-CoV elleni széleskörűen semlegesítő antitestek
csak marginális hatékonysággal rendelkeznek az olyan újonnan megjelenő
SARS-szerű CoV-törzsekkel szemben, mint az SHC014.
2. ábra: A SARS-CoV monoklonális antitestek marginális hatékonysággal rendelkeznek a SARS-szerű CoV-k ellen.
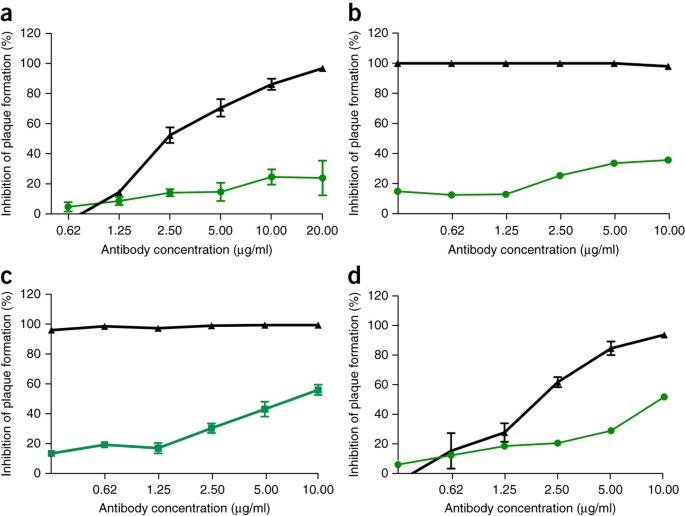
(a-d) A
monoklonális antitestek egy csoportjának - amelyek mindegyike
eredetileg járványos SARS-CoV ellen készült - hatékonyságát (a plakkok
számának csökkenéseként mérve) értékelő neutralizációs vizsgálatok Vero
sejtek SARS-CoV Urbani (fekete) vagy SHC014-MA15 (zöld) fertőzése
ellen. A vizsgált antitestek a következők voltak: fm6 (n = 3 az Urbani
esetében; n = 5 az SHC014-MA15 esetében)11,12 (a), 230,15 (n = 3 az
Urbani esetében; n = 2 az SHC014-MA15 esetében) (b), 227,15 (n = 3 az
Urbani esetében; n = 5 az SHC014-MA15 esetében) (c) és 109,8 (n = 3 az
Urbani esetében; n = 2 az SHC014-MA15 esetében)13 (d). Minden adatpont a
csoportátlagot jelenti, a hibasávok pedig az s.e.m.-t. Megjegyzendő,
hogy a b,c-ben szereplő SARS-CoV Urbani-fertőzött Vero-sejtek hibasávjai
átfedésben vannak a szimbólumokkal, és nem láthatóak.
A
meglévő vakcinák SHC014-MA15 fertőzéssel szembeni hatékonyságának
értékelésére idős egereket vakcináztunk kétszeresen inaktivált teljes
SARS-CoV-val (DIV). Korábbi munkák kimutatták, hogy a DIV képes
semlegesíteni és megvédeni a fiatal egereket a homológ vírussal való
megfertőzéssel szemben14; a vakcina azonban nem védte meg az idős
állatokat, amelyeknél fokozott immunpatológiát is megfigyeltek, ami arra
utal, hogy a vakcinázás miatt az állatok károsodhatnak15. Itt azt
találtuk, hogy a DIV nem nyújtott védelmet az SHC014-MA15 elleni
kihívással szemben a testsúlycsökkenés vagy a vírustiter tekintetében
(5a,b. kiegészítő ábra). Összhangban egy korábbi, más heterológ
csoportok 2b CoV-ival15 készített jelentéssel, a DIV-vel oltott, idős
egerekből származó szérum szintén nem semlegesítette az SHC014-MA15-öt
(5c. kiegészítő ábra).
Figyelemre
méltó, hogy a DIV-vakcináció robusztus immunpatológiát (4. kiegészítő
táblázat) és eozinofíliát eredményezett (5d-f. kiegészítő ábra). Ezek
az eredmények együttesen megerősítik, hogy a DIV vakcina nem nyújt
védelmet az SHC014 fertőzéssel szemben, és esetleg fokozhatja a
betegséget az idős vakcinázott csoportban.
Az
egerek DIV-vel történő vakcinázásával ellentétben az SHC014-MA15 élő,
attenuált vakcinaként történő alkalmazása potenciális keresztvédelmet
mutatott a SARS-CoV-vel való kihívással szemben, de az eredményeknek
fontos fenntartásai vannak. Fiatal egereket fertőztünk 104 p.f.u.
SHC014-MA15-tel, és 28 d-ig megfigyeltük őket. Ezt követően a 29. napon
SARS-MA15-tel kihívtuk az egereket (6a. kiegészítő ábra). Az egerek
előzetes fertőzése a nagy dózisú SHC014-MA15-tel védelmet nyújtott a
SARS-MA15 halálos dózisával való kihívással szemben, bár az SHC014-MA15
fertőzés után 28 nappal kiváltott antiszérákból csak minimális
SARS-CoV-neutralizációs választ kaptunk (6b. kiegészítő ábra, 1:200).
Másodlagos antigénlökés hiányában a 28 d.p.i. jelenti az
antitesttiterek várható csúcspontját, és azt jelenti, hogy idővel
csökken a SARS-CoV elleni védelem16,17. Hasonló eredményeket mutattak ki
a SARS-CoV halálos dózisával szembeni védelmet idős BALB/c egereknél a
testsúlycsökkenés és a vírusszaporodás tekintetében (6c,d. kiegészítő
ábra). A 104 p.f.u. SHC014-MA15 fertőzési dózis azonban néhány idős
állatban >10%-os súlycsökkenést és letalitást idézett elő (1. ábra és
3. kiegészítő ábra). Megállapítottuk, hogy az SHC014-MA15 alacsonyabb
dózisú (100 p.f.u.) vakcinázása nem okozott súlyvesztést, de nem védte
meg az idős állatokat a SARS-MA15 halálos dózisú kihívással szemben
(6e,f kiegészítő ábra). Az adatok együttesen arra utalnak, hogy az
SHC014-MA15 kihívás konzervált epitópokon keresztül keresztvédelmet
nyújthat a SARS-CoV ellen, de a szükséges dózis patogenezist indukál, és
kizárja a gyengített vakcinaként való felhasználást.
Miután
megállapítottuk, hogy az SHC014 tüske képes emberi sejtek fertőzését
közvetíteni és egerekben megbetegedést okozni, ezután szintetizáltunk
egy teljes hosszúságú SHC014-CoV fertőző klónt a SARS-CoV esetében
alkalmazott megközelítés alapján (3a. ábra)2 . A Vero sejtekben történő
reprodukció nem mutatott hiányt az SHC014-CoV esetében a SARS-CoV-hoz
képest (3b. ábra); azonban az SHC014-CoV szignifikánsan (P < 0,01)
gyengült primer HAE-kultúrákban a fertőzést követő 24 és 48 órában (3c.
ábra). Az egerek in vivo fertőzése nem mutatott jelentős
súlycsökkenést, de a teljes hosszúságú SHC014-CoV fertőzéssel fertőzött
tüdőben csökkent vírusreplikációt mutatott a SARS-CoV Urbani-hoz képest
(3d. ábra, e). Az eredmények együttesen igazolják a teljes hosszúságú
SHC014-CoV életképességét, de azt sugallják, hogy további adaptációra
van szükség ahhoz, hogy a replikációja egyenértékű legyen a járványos
SARS-CoV-éval emberi légzőszervi sejtekben és egerekben.
3. ábra: A teljes hosszúságú SHC014-CoV szaporodik emberi légutakban, de nem rendelkezik a járványos SARS-CoV virulenciájával.
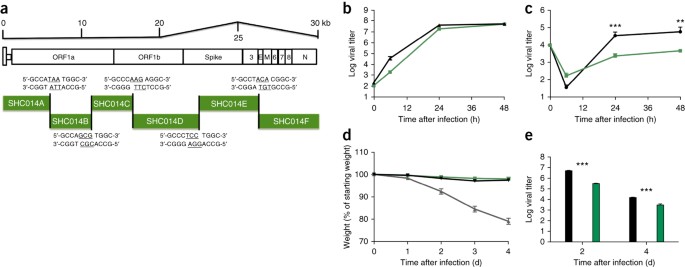
(a) Az
SHC014-CoV molekuláris klón vázlata, amelyet hat összefüggő cDNS-ként
szintetizáltak (SHC014A, SHC014B, SHC014C, SHC014D, SHC0144E és
SHC014F), amelyeket egyedi BglI-helyek szegélyeznek, amelyek lehetővé
tették a teljes hosszúságú cDNS nyílt olvasókereteket (az 1a, 1b,
spike, 3, burok, mátrix, 6-8 és nukleokapszid) kifejező cDNS irányított
összeállítását. Az aláhúzott nukleotidok a restrikciós enzim hasítása
után kialakult overhang szekvenciákat jelölik. (b,c) A SARS-CoV Urbani
(fekete) vagy az SHC014-CoV (zöld) vírusreplikációja Vero sejtek (b)
vagy jól differenciált, primer levegő-folyadék határfelületű HAE
sejttenyészetek (c) 0,01-es MOI mellett történő fertőzése után. A
mintákat egyedi időpontokban gyűjtöttük, minden csoport esetében
biológiai ismétlésekkel (n = 3). Az adatok egy kísérletet képviselnek
mind a Vero, mind a HAE sejtek esetében. (d,e) SARS-CoV MA15 esetében n =
3, SHC014-CoV esetében n = 7; SARS-Urbani esetében n = 6) (d) 10 hetes
BALB/c egerek súlyvesztése (n = 3 SARS-CoV MA15, n = 7 SHC014-CoV; n =
6 SARS-Urbani) (d) és vírusreplikáció a tüdőben (n = 3 SARS-Urbani és
SHC014-CoV) (e) 10 hetes BALB/c egereknél, amelyeket 1 × 105 p.
fertőzött. f.u. SARS-CoV MA15 (szürke), SHC014-CoV (zöld) vagy SARS-CoV
Urbani (fekete) vírussal i.n. úton. Minden adatpont a csoportátlagot
jelenti, a hibasávok pedig az s.e.m. értéket határozzák meg. **P <
0,01 és ***P < 0,001 az egyes időpontok kétfarkú Student's t-próbája
alapján.
A SARS-CoV-járvány idején gyorsan kapcsolatot találtak a pálmacibet és az emberekben kimutatott CoV-törzsek között4.
Erre a
megállapításra építve a közös megjelenési paradigma szerint a járványos
SARS-CoV denevérvírusként indult, átugrott a cibetvírusokra, és a
receptor-kötő domén (RBD) módosításait beépítette a cibetvírus Ace2-hez
való jobb kötődés érdekében (hivatkozás 18).
Az
élőállat-piacokon az emberekkel való későbbi érintkezés lehetővé tette
az emberi fertőzést a cibet törzzsel, amely viszont a járványos törzzé
vált (4a. ábra).
A
filogenetikai elemzés azonban arra utal, hogy a korai emberi
SARS-törzsek közelebbi rokonságban állnak a denevértörzsekkel, mint a
cibet törzsekkel18.
Ezért
egy másik paradigma szerint a SARS-CoV megjelenését a denevér-ember
közötti közvetlen átvitel indította el, a pálmacibet pedig másodlagos
gazdaként és rezervoárként szolgált a fertőzés folytatásához (4b.
ábra)19 . Mindkét paradigma esetében a másodlagos gazdaszervezetben a
tüskék adaptációja szükségszerűnek tekinthető, és a legtöbb mutáció
várhatóan az RBD-n belül történik, ami megkönnyíti a jobb fertőzést.
Mindkét elmélet azt feltételezi, hogy a denevér CoV-ok pooljai
korlátozottak, és hogy a gazdaszervezetre terjedő mutációk
véletlenszerűek és ritkák, ami csökkenti az emberben történő jövőbeli
megjelenés valószínűségét.
4. ábra: A koronavírusok megjelenési paradigmái.
A
koronavírustörzseket a denevérpopulációkban keringő kvázi-fajok tartják
fenn. (a,b) A hagyományos SARS-CoV megjelenési elméletek szerint a
gazdanövénytartományban előforduló mutánsok (piros kör) véletlenszerű
és ritka előfordulásokat jelentenek, amelyek lehetővé teszik alternatív
gazdák fertőzését. A másodlagos gazdaszervezet paradigma (a) szerint
egy nem emberi gazdaszervezetet megfertőz egy denevérprogenitor vírus,
és az adaptáció révén megkönnyíti az emberre történő átvitelt; az ezt
követő szaporodás az emberben a járványos vírustörzs kialakulásához
vezet. A közvetlen paradigma (b) szerint az átvitel a denevérek és az
emberek között köztes gazda nélkül történik; ezután az emberi
populációban szelekció történik a közeli rokon vírusokkal, amelyek egy
másodlagos gazdában szaporodnak, lehetővé téve a vírus folyamatos
fennmaradását és alkalmazkodását mindkettőben. (c) A kiméra SARS-szerű
vírusok adatai amellett érvelnek, hogy a kvázi fajkészletek több olyan
vírust tartanak fenn, amelyek mutációk nélkül képesek megfertőzni az
emberi sejteket (piros körök). Bár a járványok kialakulásához szükség
lehet a másodlagos vagy emberi gazdaszervezetekben történő adaptációra,
ha az SHC014 tüskét tartalmazó vírusok virulens CoV gerinccel
rekombinálódnak (zöld körvonallal jelölt körök), akkor az emberekben
járványos megbetegedés lehet az eredmény. A meglévő adatok mindhárom
paradigma elemeit alátámasztják.
Bár
tanulmányunk nem érvényteleníti a többi megjelenési útvonalat, egy
harmadik paradigma mellett érvel, amelyben a cirkuláló denevér
CoV-állományok "készenlétben" tartják a tüskés fehérjéket, amelyek
mutáció vagy adaptáció nélkül képesek megfertőzni az embert (4c. ábra).
Ezt a hipotézist illusztrálja az SHC014 tüskét SARS-CoV gerincben
tartalmazó kiméra vírus azon képessége, hogy RBD-adaptáció nélkül is
képes robusztus fertőzést okozni emberi légúti kultúrákban és egerekben.
A
korábban azonosított patogén CoV-hátterek3,20 megfigyelésével együtt
eredményeink arra utalnak, hogy a SARS-szerű újonnan megjelenő
törzsekhez szükséges kiindulási anyagok jelenleg az állati
rezervoárokban keringenek. Bár a teljes hosszúságú SHC014-CoV
valószínűleg további gerincadaptációt igényel ahhoz, hogy emberi
betegséget közvetítsen, a CoV-családokban dokumentált nagy gyakoriságú
rekombinációs események aláhúzzák a jövőbeli megjelenés lehetőségét és a
további felkészülés szükségességét.
Eddig
az állatpopulációk genomikai szűrését elsősorban új vírusok
azonosítására használták járványkitörésekben21. Az itt alkalmazott
megközelítés kiterjeszti ezeket az adatkészleteket a vírusok
megjelenésének és terápiás hatékonyságának kérdéseinek vizsgálatára. Az
SHC014 tüskével rendelkező vírusokat potenciális fenyegetésnek
tekintjük, mivel képesek szaporodni az elsődleges emberi légúti
kultúrákban, az emberi betegség legjobb rendelkezésre álló modelljében.
Ezenkívül az egerekben megfigyelt patogenezis azt jelzi, hogy az
SHC014-tartalmú vírusok képesek betegséget okozni emlős modellekben, RBD
adaptáció nélkül.
Különösen
a tüdőben a SARS-MA15-höz képest eltérő tropizmus és a teljes
hosszúságú SHC014-CoV gyengülése HAE-kultúrákban a SARS-CoV Urbanihez
képest arra utal, hogy az ACE2-kötődésen kívüli tényezők - beleértve a
tüskék processzivitását, a receptor biológiai hozzáférhetőségét vagy a
gazdaszervezet immunválaszainak antagonizmusát - hozzájárulhatnak a
kialakuláshoz. Azonban további vizsgálatokra van szükség főemlősökön
ahhoz, hogy ezeket az eredményeket az emberekre vonatkozó patogén
potenciálra fordíthassuk.
Fontos,
hogy a rendelkezésre álló terápiák sikertelensége kritikus
szükségletet határoz meg a további vizsgálatok és a kezelések
kifejlesztése szempontjából. Ezen ismeretek birtokában olyan felügyeleti
programok, diagnosztikai reagensek és hatékony kezelések állíthatók
elő, amelyek védelmet nyújtanak a 2b csoport-specifikus CoV-k, például
az SHC014 megjelenésével szemben, és ezeket más, hasonlóan heterogén
állományt fenntartó CoV-ágakra is alkalmazni lehet.
Amellett,
hogy felkészülést kínál a jövőben megjelenő vírusok ellen, ezt a
megközelítést az amerikai kormány által előírt, a funkciógyarapodási
vizsgálatokra vonatkozó szüneteltetéssel22 összefüggésben is figyelembe
kell venni.
A
korábbi megjelenési modellek alapján (4a. ábra, b. ábra) az
SHC014-MA15-hez hasonló kiméra vírusok létrehozásától nem várták a
patogenitás növekedését. Bár az SHC014-MA15 attenuált a szülői, egérre
adaptált SARS-CoV-hoz képest, hasonló vizsgálatok, amelyek a vad típusú
Urbani tüskét tartalmazó CoV-ok patogenitását vizsgálták az MA15
gerincen belül, nem mutattak súlycsökkenést egerekben és csökkent
vírusreplikációt23. Így az Urbani spike-MA15 CoV-hoz képest az
SHC014-MA15 ismét patogenezist mutat (1. ábra).
Ezen
eredmények alapján a tudományos felülvizsgálati testületek a hasonló,
keringő törzseken alapuló kiméra vírusokat építő vizsgálatokat túl
kockázatosnak ítélhetik, mivel nem zárható ki a fokozott patogenitás
emlős modellekben.
Az
egérre adaptált törzsekre vonatkozó korlátozásokkal és a menekülő
mutánsokat használó monoklonális antitestek fejlesztésével együtt a
CoV-k megjelenésének és terápiás hatékonyságának kutatása a jövőben
erősen korlátozott lehet. Ezek az adatok és korlátozások együttesen a
GOF-kutatással kapcsolatos aggályok kereszteződését jelentik; a jövőbeli
járványokra való felkészülés és azok enyhítésének lehetőségét
mérlegelni kell a még veszélyesebb kórokozók létrehozásának kockázatával
szemben. A jövőre vonatkozó politikák kidolgozásakor fontos mérlegelni
az e vizsgálatok által generált adatok értékét, és azt, hogy az ilyen
típusú kiméra vírus vizsgálatok indokolják-e a további vizsgálatokat a
velük járó kockázatokkal szemben.
Összességében
megközelítésünk a metagenomikai adatok felhasználásával azonosította a
cirkuláló denevér SARS-szerű CoV SHC014 által jelentett potenciális
veszélyt. Mivel a kimérikus SHC014 vírusok képesek emberi légúti
kultúrákban szaporodni, in vivo patogenezist okozni és a jelenlegi
terápiás szerek elől menekülni, mind a felügyeletre, mind a keringő
SARS-szerű vírusok elleni jobb terápiára szükség van. Megközelítésünk a
metagenomikai adatok felhasználását is lehetővé teszi a vírusok
megjelenésének előrejelzésére, valamint ezen ismeretek alkalmazására a
jövőben megjelenő vírusfertőzések kezelésére való felkészülésben.
Módszerek
Vírusok, sejtek, in vitro fertőzés és plakkvizsgálatok
A
vad típusú SARS-CoV-ot (Urbani), az egérhez adaptált SARS-CoV-ot
(MA15) és a kiméra SARS-szerű CoV-okat Vero E6 sejteken tenyésztettük
(az Egyesült Államok Hadsereg Orvosi Kutatóintézetének Fertőző
Betegségek Intézetétől kaptuk), Dulbecco's modified Eagle's mediumban
(DMEM) (Gibco, CA) és 5% magzati klónszérumban (FCS) (Hyclone, South
Logan, UT), valamint antibiotikummal/antimikotikummal (Gibco, Carlsbad,
CA) tenyésztettük. Az ACE2 ortológját expresszáló DBT sejteket (Baric
laboratórium, forrás ismeretlen) korábban már leírták mind az ember,
mind a cibet esetében; a denevér Ace2 szekvenciája a Rhinolophus
leschenaultiból származó szekvencián alapult, és a denevér Ace2-t
expresszáló DBT sejteket a korábban leírt módon hoztuk létre8.
Az
áltipizálási kísérletek hasonlóak voltak a korábban leírtak szerint10
előállított HIV-alapú álvírus felhasználásával végzett kísérletekhez,
amelyeket az ACE2 ortológot expresszáló HeLa sejteken (Wuhan Institute
of Virology) vizsgáltunk. A HeLa sejteket a korábban leírtak szerint
10% FCS-vel (Gibco, CA) kiegészített minimál esszenciális
tápfolyadékban (MEM) (Gibco, CA) tenyésztettük24.
A Vero
E6, DBT, Calu-3 2B4 és primer humán légúti epitélsejtekben a korábban
leírtak szerint végeztük el a növekedési görbéket8,25 . A munka
sejtvonal állományok egyikét sem hitelesítették vagy vizsgálták a
közelmúltban mikoplazma szempontjából, bár a munka állományok
létrehozásához használt eredeti magállományok mentesek a
szennyeződéstől. A HAE-kultúrákhoz használt emberi tüdőt az
Észak-Karolinai Egyetem Chapel Hill-i intézményi felülvizsgálati
bizottsága által jóváhagyott protokollok szerint szereztük be. A
HAE-kultúrák magasan differenciált emberi légúti epithelt képviselnek,
amely ciliált és nem ciliált epithelsejteket, valamint goblet sejteket
tartalmaz. A tenyészeteket felhasználás előtt több hétig levegő-folyadék
határfelületen is növesztik, a korábban leírtak szerint26.
Röviden,
a sejteket PBS-szel mostuk, majd beoltottuk a vírussal vagy PBS-ben 40
percig 37 °C-on mock-hígított vírussal. A beoltás után a sejteket
háromszor mostuk, és friss tápfolyadékot adtunk hozzá, hogy a "0"
időpontot jelezzük. Minden egyes leírt időpontban három vagy több
biológiai ismétlést gyűjtöttünk be. A mintagyűjtések során nem
alkalmaztunk vakítást, és a mintákat nem randomizáltuk. Minden
vírustenyésztést egy 3. biológiai biztonsági szintű (BSL)
laboratóriumban végeztünk, a biológiai biztonsági szekrényekben
redundáns ventilátorokkal, ahogyan azt csoportunk korábban leírta2. A
személyzet minden tagja motoros légtisztító légzőkészüléket (Breathe
Easy, 3M) viselt Tyvek-öltözékkel, kötényt és bakancsot, és dupla
kesztyűt viselt.
Szekvencia klaszterezés és strukturális modellezés.
A
reprezentatív CoV-k tüskéinek teljes hosszúságú genomi szekvenciáit és
az S1 domének aminosav szekvenciáit a Genbankból vagy a Pathosystems
Resource Integration Centerből (PATRIC) töltöttük le, a ClustalX
segítségével illesztettük össze, és filogenetikai összehasonlítást
végeztünk maximum likelihood becsléssel 100 bootstrap segítségével,
illetve a PhyML csomag segítségével. A fát a PhyML csomag segítségével,
maximális valószínűséggel generáltuk. A skálasáv a nukleotidcseréket
jelöli. Csak a 70% feletti bootstrap-támogatással rendelkező csomópontok
vannak jelölve.
A fa
azt mutatja, hogy a CoV-k három különböző filogenetikai csoportra
oszlanak, amelyeket α-CoV-ként, β-CoV-ként és γ-CoV-ként definiáltak. A
klasszikus alcsoportok klasztereit a β-CoV-ok esetében a 2a, 2b, 2c és
2d, az α-CoV-ok esetében pedig az 1a és 1b jelöli. A szerkezeti
modelleket a Modeller (Max Planck Institute Bioinformatics Toolkit)
segítségével generáltuk a SARS RBD SHC014 és Rs3367
homológiamodelljeihez az ACE2-vel alkotott komplexben a 2AJF
kristályszerkezet (Protein Data Bank) alapján. A homológiamodelleket
MacPyMol (1.3-as verzió) programban vizualizáltuk és manipuláltuk.
SARS-szerű kiméra vírusok előállítása.
Mind a
vad típusú, mind a kiméra vírusokat a SARS-CoV Urbani vagy a megfelelő
egérhez adaptált (SARS-CoV MA15) fertőző klónból (ic) származtattuk a
korábban leírtak szerint27.
Az
SHC014 tüskeszekvenciáit tartalmazó plazmidokat restrikciós emésztéssel
kivontuk, és az MA15 fertőző klón E és F plazmidjába ligáltuk. A klónt
hat összefüggő cDNS-ként tervezték és vásárolták a Bio Basic-tól, a
publikált szekvenciák felhasználásával, amelyeket egyedi II. osztályú
restrikciós endonukleáz-helyek (BglI) szegélyeznek. Ezt követően a vad
típusú, kiméra SARS-CoV és SHC014-CoV genomrészleteket tartalmazó
plazmidokat felszaporítottuk, kivágtuk, ligáltuk és tisztítottuk.
Ezután
in vitro transzkripciós reakciókat végeztünk a teljes hosszúságú genomi
RNS szintéziséhez, amelyet a korábban leírtak szerint Vero E6 sejtekbe
transzfektáltunk2. A transzfektált sejtekből származó tápfolyadékot
leszedtük, és az a későbbi kísérletekhez magkészletként szolgált. A
kiméra és a teljes hosszúságú vírusokat szekvenciaelemzéssel igazoltuk,
mielőtt e vizsgálatokban felhasználtuk volna. A kimérikus mutáns és
teljes hosszúságú SHC014-CoV szintetikus konstrukcióját az
Észak-Karolinai Egyetem Intézményi Biológiai Biztonsági Bizottsága és a
Kettős felhasználású, aggodalomra okot adó kutatások bizottsága hagyta
jóvá.
Etikai nyilatkozat
Ezt a
vizsgálatot az NIH laboratóriumi állatok gondozására és felhasználására
vonatkozó, az Office of Laboratory Animal Welfare (OLAW) által kiadott
ajánlásoknak megfelelően végeztük. A Chapel Hill-i Észak-Karolinai
Egyetem (UNC, engedélyszám: A-3410-01) intézményi állatgondozási és
-használati bizottsága (IACUC) jóváhagyta az e vizsgálatokban
alkalmazott állatkísérleti protokollt (IACUC #13-033).
Egerek és in vivo fertőzés
Nőstény,
10 hetes és 12 hónapos BALB/cAnNHsD egereket a Harlan Laboratories-tól
rendeltünk. Az egérfertőzéseket a korábban leírtak szerint végeztük20.
Röviden, az állatokat egy BSL3 laboratóriumba vitték, és a fertőzés
előtt 1 hétig hagyták őket akklimatizálódni. A fertőzéshez és az élő,
hígított vírusvakcinációhoz az egereket ketamin és xilazin keverékével
altatták, és intranazálisan fertőzték meg őket, amikor kihívták őket,
50 μl foszfát-pufferelt sóoldattal (PBS) vagy hígított vírussal, három
vagy négy egérrel, időpontonként, fertőzési csoportonként és
dózisonként, az ábra legendáiban leírtak szerint.
Az
egyes egerek esetében a fertőzésre vonatkozó megjegyzések, beleértve a
teljes adag belégzésének elmulasztását, az inokulum orrból történő
buborékosodását vagy a szájon keresztül történő fertőzést, a kutató
döntése alapján az egéradatok kizárásához vezethettek; a fertőzés után
nem határoztak meg más, előre meghatározott kizárási vagy felvételi
kritériumokat. Az állatkísérletekben nem alkalmaztak vakítást, és az
állatokat nem randomizálták. A vakcinázáshoz a fiatal és idős egereket
lábpárna-injekcióval vakcinázták 20 μl térfogatú, 0,2 μg kettős
inaktivált SARS-CoV vakcinával, alumíniummal vagy PBS-sel; az egereket
22 nappal később ugyanezzel a kezelési móddal fokozták, majd 21 nappal
később kihívták őket.
Minden
csoport esetében, a protokoll szerint, az állatokat a kísérlet
időtartama alatt naponta megfigyelték a betegség klinikai tünetei
(púposodás, felborzolt szőrzet és csökkent aktivitás) szempontjából. A
súlyvesztést az első 7 napban naponta ellenőrizték, majd a
testtömeg-ellenőrzést addig folytatták, amíg az állatok vissza nem
álltak a kezdeti kiindulási súlyukra, vagy 3 napig folyamatosan
súlygyarapodást mutattak.
Minden
olyan egeret, amely a kiindulási testsúlyának 20%-ánál nagyobb
mértékben veszített, daráltak és naponta többször is megfigyeltek, amíg
a 20%-os határérték alatt maradtak. Azokat az egereket, amelyek kezdeti
testsúlyuk 30%-ánál nagyobb mértékben veszítettek, a protokoll szerint
azonnal feláldozták. Minden olyan egeret, amelyről úgy ítélték meg,
hogy haldoklik vagy valószínűleg nem fog felépülni, a kutató döntése
alapján szintén humánusan feláldozták. Az eutanáziát izoflurán
túladagolással végeztük, és a halál beálltát nyaki dislokációval
igazoltuk. Minden egérvizsgálatot az Észak-Karolinai Egyetemen végeztek
(Animal Welfare Assurance #A3410-01) az UNC intézményi állatgondozási
és -használati bizottsága (IACUC) által jóváhagyott protokollok
alapján.
Hisztológiai elemzés.
A bal
tüdőt eltávolítottuk, és 1 hétig 10%-os pufferelt formalinban (Fisher),
felfúvás nélkül tartottuk. A szöveteket paraffinba ágyazták, és az UNC
Lineberger Comprehensive Cancer Center szövettani központja 5 mm-es
metszeteket készített. Az antigénfestés mértékének meghatározásához a
metszeteket a vírusantigénre festettük a kereskedelmi forgalomban
kapható poliklonális SARS-CoV anti-nukleokapszid antitesttel (Imgenex),
és vakon pontoztuk a légutak és a parenchima festődését a korábban
leírtak szerint20. A képeket Olympus DP71 kamerával felszerelt Olympus
BX41 mikroszkóppal készítettük.
Vírusneutralizációs vizsgálatok.
A
plakkredukciós semlegesítési titerpróbákat korábban jellemzett SARS-CoV
elleni antitestekkel végeztük, a korábban leírtak szerint11,
12,13
.
Röviden, a neutralizáló antitesteket vagy szérumot sorozatban
kétszeresére hígítottuk, és a különböző fertőző klónok, SARS-CoV törzsek
100 p.f.u.-jával inkubáltuk 1 órán keresztül 37 °C-on. A vírust és az
antitesteket ezután 5 × 105 Vero E6 sejtet/lyuk tartalmazó 6 lyukú
lemezbe adtuk, többszörös replikációval (n ≥ 2). Az 1 órás 37 °C-on
történő inkubációt követően a sejteket 3 ml 0,8%-os agarózzal fedtük le a
táptalajban. A lemezeket 2 napig 37 °C-on inkubáltuk, majd 3 órán át
semleges vörössel festettük, és megszámoltuk a plakkokat. A plakkok
csökkenésének százalékos arányát a következőképpen számoltuk ki: (1 -
(antitesttel rendelkező plakkok száma/antitest nélküli plakkok száma)) ×
100.
Statisztikai elemzés
Minden
kísérletet két kísérleti csoport (vagy két vírus, vagy vakcinázott és
nem vakcinázott kohorsz) szembeállításával végeztünk. Ezért a
vírustiter és a szövettani pontszámok szignifikáns különbségeit az
egyes időpontokban kétfarkú Student's t-próbával határoztuk meg. Az
adatok normális eloszlásúak voltak minden összehasonlított csoportban,
és hasonló szórással rendelkeztek.
Biológiai biztonság és biológiai védelem
A
bejelentett vizsgálatokat azután kezdtük el, hogy az Észak-Karolinai
Egyetem Intézményi Biológiai Biztonság Bizottság jóváhagyta a kísérleti
protokollt (Project Title: A denevér SARS-szerű CoV-k fertőző
klónjainak előállítása; Laborbiztonsági terv azonosítója: 20145741;
G-terv azonosítója: 12279).
Ezeket a vizsgálatokat az USA
kormányának az influenza-, MERS- és SARS-vírusokat érintő, a
funkciónyereséggel kapcsolatos kiválasztott kutatásokról szóló,
tanácskozási eljárás keretében történő kutatási finanszírozás
szüneteltetése előtt kezdték meg. Ezt a dokumentumot a finanszírozó
ügynökség, az NIH felülvizsgálta. A vizsgálatok folytatását kérték, és
ezt az NIH jóváhagyta.
A
SARS-CoV szelektív kórokozó. E tanulmányok minden munkáját a SARS-CoV,
MERs-CoV és más rokon CoV vírusokra jóváhagyott szabványos működési
eljárások (SOP) és biztonsági feltételek szerint végezték. Intézményi
CoV BSL3 létesítményeinket úgy alakítottuk ki, hogy megfeleljenek a
Biosafety in Microbiological and Biomedical Laboratories (BMBL), az
Egyesült Államok Egészségügyi és Humán Szolgálatának, a Közegészségügyi
Szolgálatnak, a Centers for Disease Control (CDC) és az NIH által
ajánlott biztonsági követelményeknek. A laboratóriumi biztonsági
terveket benyújtották az UNC Környezeti Egészségügyi és Biztonsági
Osztályának (EHS) és a CDC-nek, és a létesítményt jóváhagyták. A
létesítménybe való belépéshez elektronikus kártyás belépésre van
szükség.
Az EHS
minden dolgozót kiképzett a motoros légtisztító légzőkészülék (PAPR)
biztonságos használatára, és a BSL3 létesítményben megfelelő
munkaszokások, valamint aktív egészségügyi felügyeleti tervek vannak
érvényben. A CoV BSL3 létesítményeink redundáns ventilátorokat,
vészhelyzeti áramellátást tartalmaznak a ventilátorokhoz és a biológiai
biztonsági szekrényekhez és fagyasztókhoz, és létesítményeinkben
SealSafe egérállványok is elhelyezhetők. A BSL3 kórokozóként besorolt
anyagok közé tartoznak a SARS-CoV, a denevér CoV prekurzor törzsek, a
MERS-CoV és az ezekből a kórokozókból származó mutánsok. A BSL3
létesítményekben a fertőző vírussal végzett kísérleteket II. osztályú,
tanúsított biológiai biztonsági szekrényben (BSC) végzik.
A
személyzet minden tagja műtősruhát, Tyvek-öltözetet és kötényt, PAPR-t
és cipőtakarót visel, és kezeiket dupla kesztyűvel védik. A
BSL3-felhasználókra az Egyetemi Alkalmazottak Foglalkozás-egészségügyi
Klinikája (UEOHC) által felügyelt egészségügyi felügyeleti terv
vonatkozik, amely magában foglalja az éves orvosi vizsgálatot, az éves
influenza elleni védőoltást és a CoV-fertőzéssel kapcsolatos tünetek
kötelező bejelentését a BSL3-ban való munkavégzés idején. Minden
BSL3-felhasználót kiképeztek az expozíció kezelésére és a jelentési
protokollokra, felkészültek az önkaranténra, és képzést kaptak arra,
hogy vészhelyzetben biztonságosan eljuttassák a helyi fertőző
betegségekkel foglalkozó osztályra. Az EHS és az UEOHC minden lehetséges
expozíciós eseményt jelent és kivizsgál, és a jelentéseket a CDC-nek
és az NIH-nek is benyújtja.
Referenciák letöltése
https://pokolafoldon.hu/hihetetlen/betiltva-covid-eszak-karolinaban-kezdodott-nem-wuhanban